Detection and processing of images
Click on the link to Eye Design Book to see various pictures of the structure of the eye and retina.
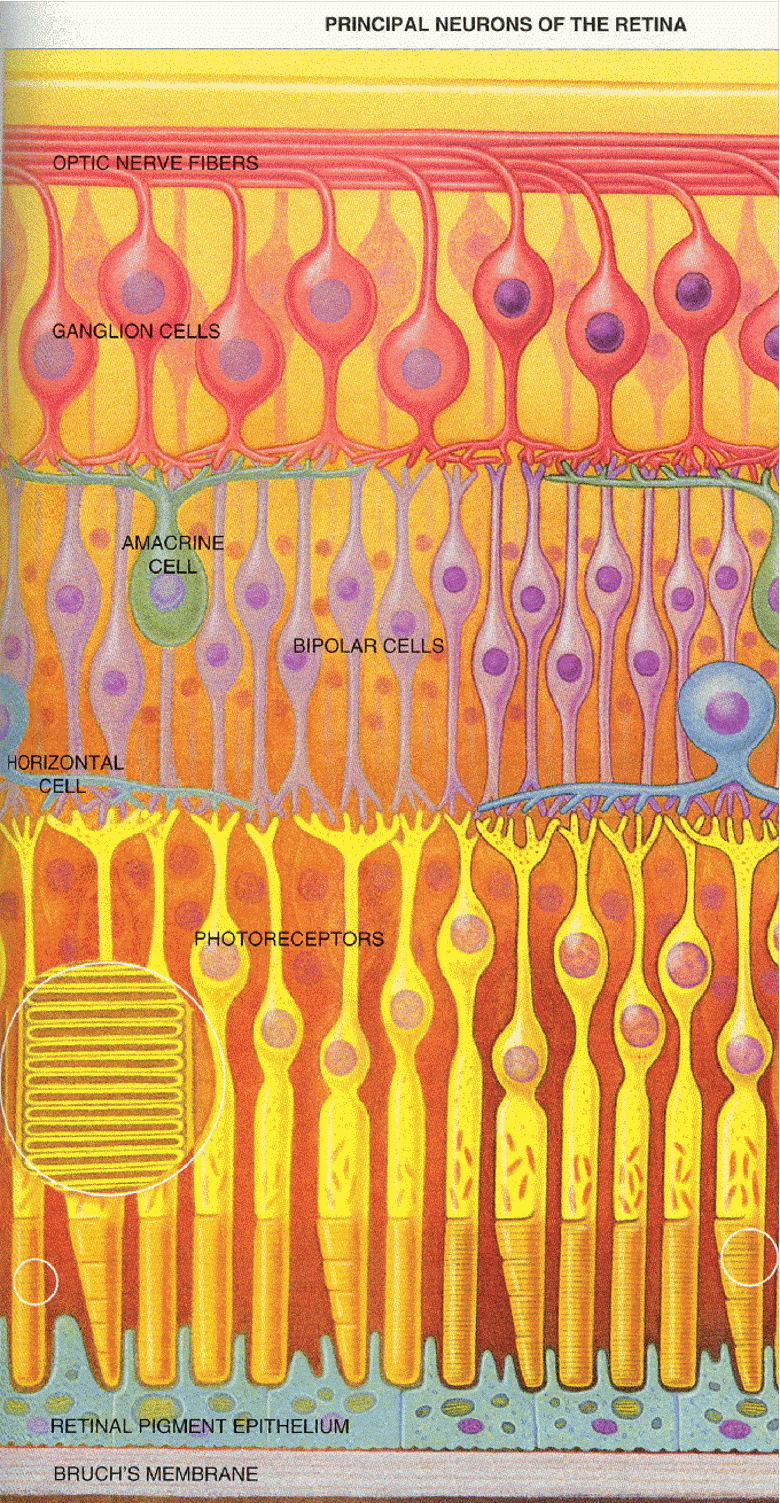
The retina is a thin layer of cells that sits at the back of the eye. It is covered with small blood vessels, but because these are always present (and therefore do not move relative to the retina) they are generally invisible. Behind the vessels are several layers of different types of nerve cells (also known as neurons). Moving from the front of the eye to the back the last layer of cells we encounter are the light detecting cells: the rods and the cones.
Some interesting points about the retina.
The retina has a layer of blood vessels that are in the path of the light reaching the retina. Normally we never see these blood vessels even though the sit in front of the image that forms on the retina. This is because they never move relative to the retina. Images that do not move over the surface of the retina are ignored by the brain. However these blood vessels can be visualized by shining a small light into the eye and moving it around. This causes shadows of the blood vessels which move around and make an image of the vessels visible.
Remember that the image on the retina is inverted relative to the object. The brain actually flips the image over. You can tell this by gently pressing a finger against your eyelid close to your nose. This pressure briefly cuts off oxygen to the rods and cones directly behind it and they report "darkness." However notice that the brain reverses the position of the dark dot that gets created.
Finally, there is an area off the center of the eye where the axons of the optic nerve gather and leave the retina. There is a spot there, where there are no rods and cones. Look out at the world. Do you see a hole in your visual field? No. This is because the brain fills in the missing parts. However you can show that there is a blind spot in the following way: Close one eye. Hold up a finger even with your nose about one foot away. Focus your eye intently on your finger. Now hold up a pen even with your finger and gradually move it away from the center. You may have to move it up and down a little. Pretty soon you will find a location where the top of the pen or pencil disappears. This occurs when the image falls on the blind spot.
The anatomy of the retina
The retina is made up of several layers
of nerve cells (also known as neurons).
For a picture of the cell layers that make up the retina go to the following
link on the Eye
Design Book.
{kind=link}
The main cells we are interested in are (1) the rods and cones (which capture photons and respond with an electrical response), (2) the bipolar cells, which carry the signal from the rods and cones, and (3) the ganglion cells, which pick up the electrical signal and carry it to the brain. There are two other types of cells (amacrine cells and bipolar cells) which carry messages sideways along the retina and allow the other cells to interact with one another. The complexity of the retina is our first hint that the retina does more than simply pass an imprint of the optical image to the brain. A lot of neural processing goes on here. In fact the cells in the retina are similar to nerve cells in the brain, and some scientists have described the retina as an extension of the brain.
Before I go into detail about the cells in the retina I need to say a little about cells in general, and about nerve cells in particular.
CELLS
Nerve cells are an example of a type of cell called an excitable cell. All
cells have a small negative charge on the inside (relative to the body fluid
outside the cell). Excitable cells are those which can change their internal
charge.
All living things are made of cells.
Each cell consists of a membrane which surrounds it and a set of internal
organelles such as the nucleus, mitochondria etc. The membrane which is made
up of fat and protein is highly structured and controls what can get in and
out of the cell.
So, cells are basically little bags of fluid and gunk inside it surrounded
by a membrane which controls what can move in or out of a cell. Most of the
volume of a cell is water. Dissolved in this water are lots of large and small
molecules. One very important kind of molecule is called a salt. A salt consists
of a positively charged moleculed linked up with a negatively chared molecule.
Examples are NaCl (Sodium chloride, table salt) or KCl (potassium chloride).
In water the components of salts break apart into two charged molecules, known
as ions. Thus NaCl becomes Na+ and Cl-. And KCl become K+ and Cl-. These charged
ions float around inside and outside the cell. However the cell is very particular
about its ions. Special pumps in the cell membrane actively pump Na+ out of
the cell, and pump K+ into the cell. More Na+ gets pumped out than K+ gets
pumped in, and the overall result is that there is a net negative charge inside
the cell. The negative charge is actually carried by Cl- and other large negatively
charged molecules.
A cell is thus like a tiny battery. All cells have a voltage inside that is negative relative to the outside by about -70millivolts (1 millivolt - 1/000 th of a volt).
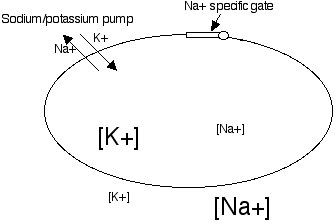
As shown in the figure above cells have
much more K+ (the symbol [K+] indicates the concentration of K+) inside the
cell than outside. There is much more Na+ outside the cell than inside.
EXCITABLE CELLS
Most cells just stay negatively charged
for their entire life. However there is one type of cell that isn't content
to stay the same charge -- these are called excitable cells. There are three
kinds: nerve cells (also known as neurons), muscle cells, and eggs. What makes
them different is that the membrane is filled with special channels which
have two important properties: they can open or close when an appropriate
stimulus occurs and the SELECTIVELY let a single type of ion pass into or
out of the cell through the membrane. For example most excitable cells have
lots of gated channels for Na+. This is illustrated in the previous figure.
When these gates open lots of Na+ goes rushing into the cell because (1) the
cell is negatively charged and (2) there is lots more Na+ outside than inside
the cell (because the cell normally pumps most of the Na+ out of the cell).
This is how nerve cells work. Special stimuli cause the Na+ gates on parts of their membranes to open. This makes the cell less negative -- we say that it depolarizes -- or may even become positively charged inside. In some types of cells this depolarization is just a local event (i.e. occurs only at one spot in the cell). In many kinds of nerve cells, there are lots of Na+ gates spread all along the surface of the cell, and what's more these Na+ gates in nerve cells open in response to a change in the voltage in the cell. In fact what opens the gates of nerve cell Na+ channels is when the cell becomes less negative. However, the effect of opening up Na+ gates is to make the cell less negative. A system in which a stimulus and effect are the same is called a POSITIVE FEEDBACK LOOP. The less negative the inside of a nerve cell become the more Na+ channels get opened --> which makes the cell even less negative.
Since opening Na+ gates causes the cell to become less negative, and there are lots of these Na+ gates which respond to reduced cell negativity all along the cell, the change in the voltage of the cell quickly spread down the cell. After a short time, however, the sodium gates close all by themselves and the cell pumps the Na+ back out. In this way a wave of depolarization can travel down the length of cell. The cell quickly "resets" and gets ready for the next wave.
Most nerve cells have long thin extensions called AXONS. When a brief wave of voltage change spread down a nerve cell's axon, it is referred to as an action potential or a nerve spike or a nerve impulse. Neurons (or nerve cells) of this type are very important in the nervous system. They can send messages long distances over long extensions (the axons). When the nerve spikes reach the end of the axon, it causes neurotransmitter release and can excite (or inhibit) any cell it is attached to.
EXCITABLE CELLS IN THE RETINA
RODS and CONES. Light travels through the clear layers of nervous tissue
in the retina and reaches the rods and cones. Rods and cones are actually
very similar. The following description is of a rod, but it applies to both
rods and cones fro the most part.
Each rod has an inner segment (includes the nucleus, mitochondria etc) and they have a foot (called the "synaptic ending"). Like all cells the entire cell is surrounded by a membrane.
The outer segment is full of stacks of membrane-enclosed discs each shaped like a pancake. Stuck in the membranes of these disks are lots of pigment molecules. A pigment is the name given to any molecule is particularly good at absorbing some wavelengths of light. In the case of the pigment in rods, it looks purple because it tends to absorb green light extremely well, and reflects or transmits some red and blue light which makes it look purple.
This pigment is known as "visual purple" or "rhodopsin". Rhodopsin has two parts -- a molecule called RETINAL (a modified version of vitamin A (the stuff found in carrots!) and a molecule called OPSIN.
When a photon of light strikes rhodopsin
and gets absorbed the energy that is incorporated into the molecule causes
a series of complex changes in the molecule's structure. This causes the retinal
and opsin to split apart and a whole series of chemical reactions get triggered
in the rod outer segment. Once a pigment molecule absorbs a photon and splits,
it can no longer absorb photons. It now no longer looks purples -- it can't
absorb any light so all wavelengths get reflected or transmitted and it looks
white.
We call a rhodopsin molecule that has absorbed a photon and split into retinal
and opsin bleached. UNBLEACHED RHODOPSIN CAN ABSORB LIGHT, BLEACHED RHODOPSIN
CAN'T. Eventually the cell will take the bleached rhodopsin and rebuild
its parts into fresh, unbleached rhodopsin, but this process takes several
minutes.
SENSITIVITY
When lots of photons are arriving at the retina many of the rhodopsin molecules get bleached, and as they get reformed that get bleached again. When a lot of photons are arriving quickly the ratio of bleached to unbleached rhodopsin gets high, and the rod is relatively bad at absorbing photons. The eye becomes less sensitive.
When there are very few photons striking the eye, the eye rebuilds all of its rhodopsin and the rods fill up with unbleached rhodopsin. Any photon entering the eye is very likely to be captured. In this state the eye is very sensitive. The figure below illustrates this:
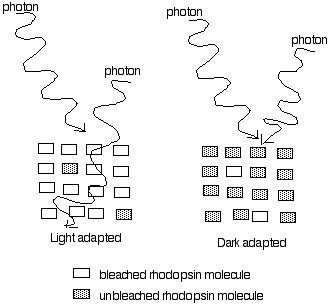
When lots of photons are striking the eye (left half of figure above) most of the rhodopsin in a rod becomes bleached. If photons do not strike unbleached rhodopsin they travel right past the retina and have no effect. Thus in bright light the retina has very little unbleached rhodopsin, and captures relatively few photons. The eye in this state is "insenstive" and responds only to large changes in light intensity.
When the eye is placed in the dark, little new bleaching of rhodopsin takes places. Gradually the density of unbleached rhodopsin builds up (right side of figure above). In this condition any photons arriving are very likely to be absorbed and cause excitation. This state is called dark adapted.
The process in which the eye becomes highly sensitive by building up lots of unbleached rhodopsin is called dark adaptation. While build-up of unbleached rhodopsin (because the eye makes more, and relatively little gets broken down) is the main factor contributing to the increase in sensitivity during dark adaptation there are other contributing factors as well, which we will discuss later.
TRANSMISSION OF THE MESSAGE
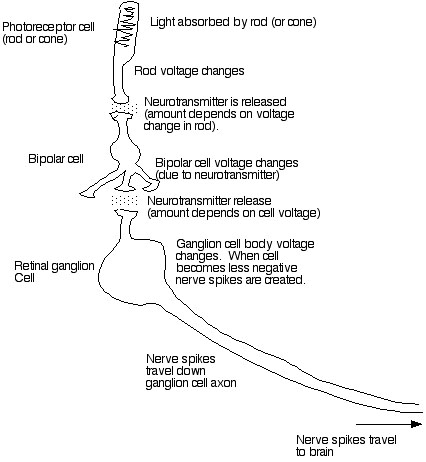
Once light strikes rhodopsin and the rhodopsin gets broken down a complex set of chemical reactions occur. The last of these reactions involves the opening up of gates which let sodium into the cell. This causes a change in the voltage within the cell. Thus rods and cones are actually excitable nerve cells. The thing that causes their voltage to change is the absorption of light by rhodopsin.
Once a rod or cone gets "excited" (i.e. its internal voltage changes) it releases a substance from its foot called a neurotransmitter. This neurotransmitter travels across a small space between the foot of the cone and the next cell in line: the BIPOLAR CELL. The small space between the cells is called a synapse. After this special neurotransmitter molecule travels across the gap between the rod (or cone) and the bipolar cell it binds to the bipolar cell. THIS CAUSES THE OPENING OF SODIUM GATES IN THE BIPOLAR CELL. The bipolar cell then becomes less negative (because Na+ moves in), and it releases neurotransmitter to the next cell in line: the GANGLION CELL. This causes depolarization of the ganglion cell.
RETINAL GANGLION cells differ from photoreceptor cells and bipolar cells in one important way. They have the special voltage-sensitive Na+ channels I described earlier, which allows them to generate ACTION POTENTIALS. These action potentials or nerve spikes travel down the length of the ganglion cell. Ganglion cells have long extensions called axons which travel all the way from the retina to the brain. The axons of the retinal ganglion cells are grouped together to form the OPTIC NERVE.
IN SUMMARY: Rods and cones capture photons of light. This causes the cells to change their internal charge. This causes release of neurotransmitter to bipolar cells. This causes them to change their charge. This causes release of neurotransmitter onto ganglion cells. This causes nerve spikes which travel out of the retina to the brain.
It is also possible for cells to "inhibit" one another. For example some bipolar cells release a different type of neurotransmitter which makes ganglion cells less likely to fire action potentials. Thus some inputs are excitatory and some are inhibitory. Retinal cells also send messages across the retina to other retinal cells (through the horizontal cells and amacrine cells which link different bipolar cells and different ganglion cells together). In this way it is possible for a single rod or cone to excite one set of ganglion cells while at the same time inhibiting some other ganglion cells nearby.
Last Modified
Tuesday, 21-Oct-2003
fleishml@union.edu